

0 引言
【研究意义】大麦在世界谷类作物中的种植面积和产量仅次于小麦,水稻和玉米,位居第四位[1-2]。大麦也是我国干旱、半干旱地区和气候偏冷凉地区重要的种植作物[3],大麦籽粒兼具食用、饲用、酿酒等用途,大麦秸秆是优质的禾谷类饲草[4]。大麦具有生育期相对较短、熟期早、耐贫瘠、适应性广及抗逆性强等特点[5-6]。我国大麦生产自给率尚不高[7-8],而新疆牲畜养殖业以及啤酒、白酒等酿造产业的发展对大麦的需求旺盛,另外新疆高海拔以及冷凉气候也决定了其成为我国优质大麦的主产区之一[9]。培育高产、优质、多抗性新品种是稳定和发展大麦产业的重要前提,育种材料遗传基础狭窄,制约了大麦品种选育。从国外引进大麦种质资源能够丰富我国的大麦育种基础,有利于选育出优良的后代材料。【前人研究进展】对引进种质资源材料进行正确合理的鉴定和评价是进行有效利用的基础[10-11]。对大麦种质资源的表型性状多样性分析与评价是一项关键的工作[12],新品种的选育及其生产发展取决于对优异资源研究的深度,大麦种质资源的鉴定评价是对其进行合理利用的前提[13-14]。了解优异大麦种质可加快育种进程,有助于丰富大麦育种亲本类型,加强优异基因的挖掘与利用,为大麦杂交育种和良种选育提供重要的依据[15-16]。【本研究切入点】目前,针对新疆大麦农艺性状遗传多样性的分析报道较少,且各研究所用的大麦种质资源材料来源不同,种植区域也不同,生态环境差异较大,因此表现出不同的遗传多样性[9]。【拟解决的关键问题】引自塔吉克斯坦、吉尔吉斯斯坦和哈萨克斯坦的276份中亚大麦种质资源为材料,对其在我国新疆的农艺及品质性状进行综合评价,分析引进的中亚大麦种质资源的遗传多样性,筛选出性状优良的大麦资源材料作为育种亲本,为我国新疆大麦育种改良和生产中品种的选择提供理论支撑。
1 材料与方法
1.1 材料
以引进于塔吉克斯坦、吉尔吉斯斯坦、哈萨克斯坦的276份大麦种质为供试材料,其中塔吉克斯坦占9.4%,吉尔吉斯斯坦占60%,哈萨克斯坦占30.6%。
1.2 方法
1.2.1 试验设计
试验于2021年4月在新疆农业科学院奇台麦类试验站(平均海拔823 m,N 44°13',E 89°12')进行,试验地土壤为黑壤土(肥力中等),前茬作物为小麦。采用随机区组设计,每份材料播种4行,行长2 m,行宽1.2 m,行距30 cm,株距2 cm,3次重复。田间管理与大田常规管理相同。
根据《大麦种质资源描述规范和数据标准》中的规定[17]田间调查记载幼苗生长习性、分蘖力、穗密度、芒性、侧小穗、穗轴茸毛、棱型等项目,成熟期随机选取长势一致,株型一致的10株植株进行室内考种,考察株高、穗长、单株穗数、每穗粒数、千粒重5个数量性状。
1.2.2 测定指标
遗传多样性指数(Shannon- Wiener diversity index,H')的计算采用 Shannon-weaver。H'=-∑Piln Pi
式中,Pi为某一性状第i个级别出现的频率。
对数量性状进行分级,对质量性状予以赋值,多样性指数的分级方法:先计算参试材料总体平均数(X)和标准差(d),然后划分为10级,从第1级[Xi<(X-2d)] 到第10级 [Xi>(X+2d)],每0.5d为1级,每一级的相对频率用于计算多样性指数[18]。
1.3 数据处理
使用 Microsoft Excel 软件对数据进行初步整理,计算各性状的最大值、最小值、平均值、极差、标准差和变异系数。利用 Origin软件采用系统聚类分析方法,以平均数进行聚类。
2 结果与分析
2.1 大麦种质资源质量性状遗传多样性
研究表明,276份大麦种质资源7个质量性状在不同材料间表现出不同程度的多样性。供试的大麦材料幼苗生长习性多为半直立型,占参试材料的57%;其次为直立型,占40%;匍匐型最少仅占3%。分蘖力以中等为主,占比为87%;穗密度以稀疏型为主,占比为82%;芒性以齿为主,占比为69%;侧小穗以无居多,占比为79%;穗轴茸毛以长为主,占比为87%;棱型以二棱大麦为主,占比为73%,其次是多棱大麦,占比为27%。
供试大麦7个质量性状之间的遗传多样性指数变幅为0.39~0.79,其中幼苗生长习性的遗传多样性指数最高(0.79),其余6个性状的遗传多样性指数从大到小依次为芒性(0.62)、棱型(0.58)、侧小穗(0.51)、分蘖力(0.48)、小穗密度(0.47)、穗轴茸毛(0.39)。表1
表1 7个质量性状的遗传多样性
Tab.1
| 性状 Traits | 遗传多样性指数(H') Genetic diversity index | 频率分布 Ratio of distribution | |||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | ||
| 幼苗生长习性 Seedling growth habit | 0.79 | 0.57 | 0.40 | 0.00 | 0.03 |
| 分蘖力 Tillering ability | 0.48 | 0.10 | 0.87 | 0.03 | |
| 穗密度 Spike density | 0.47 | 0.82 | 0.18 | ||
| 芒性 Awn character | 0.62 | 0.31 | 0.69 | ||
| 侧小穗 Lateral spikelet | 0.51 | 0.79 | 0.21 | ||
| 穗轴茸毛 Cob pubescence | 0.39 | 0.13 | 0.87 | ||
| 棱型 Prismatic type | 0.58 | 0.73 | 0.00 | 0.27 | |
注:幼苗生长习性1~4依次为半直立、直立、半匍匐、匍匐;分蘖力1~3依次为弱、中、强;穗密度1~2依次为稀、密;芒性1~2依次为光、齿;侧小穗1~2依次为无、多;穗轴茸毛1~2依次为短、长;棱型1~3依次为二棱、六棱、多棱
Note: Seedling growth habit 1-4 are semierect, upright, half-punched and creeping; tillering ability 1-3 are weak, medium, strong; spike density 1-2 are sparse, Dense; Awn character 1-2 are smooth and Serrated; Lateral spikelet1-2 are None and some; Cob pubescenc1-2 are short and long; prismatic type 1-3 are two ridges,six ridges and multi prismatic
2.2 数量性状遗传多样性
研究表明,数量性状的变异系数为0.75%~4.26%,其中每穗粒数的变异系数最大(4.26%),千粒重的变异系数最小(0.75%)。一般变异系数大于10%表示样本间差异比较大,参试材料的5个数量性状变异范围较小。
参试大麦材料5个数量性状的遗传多样性指数范围为1.57~2.03,从大到小依次为株高(2.03)>千粒重(2.02)>穗长(1.99)>单株穗数(1.98)>每穗粒数(1.57),数量性状的遗传多样性指数高于质量性状,5个数量性状在不同材料间表现出丰富的遗传多样性。表2
表2 5个数量性状的遗传多样性
Tab.2
| 性状 Traits | 最大值 Maximum | 最小值 Minimum | 极差 Range | 平均值 Mean | 标准差 Standard deviation | 变异系数 Coefficient of variation (%) | 遗传多样性指数(H') Genetic diversity index |
|---|---|---|---|---|---|---|---|
| 株高 Plant height(cm) | 104.00 | 48.00 | 56.00 | 86.50 | 8.52 | 0.99 | 2.03 |
| 千粒重 Thousand grain weight(g) | 60.20 | 40.50 | 19.70 | 51.44 | 3.84 | 0.75 | 2.02 |
| 单株穗数 Pericles per plant | 8.40 | 2.20 | 6.20 | 3.64 | 0.80 | 2.19 | 1.98 |
| 穗长 Panicle length(cm) | 14.78 | 2.88 | 11.90 | 7.26 | 1.71 | 2.36 | 1.99 |
| 每穗粒数 Grain per spike | 88.56 | 12.78 | 75.78 | 27.51 | 11.73 | 4.26 | 1.57 |
2.3 大麦种质资源数量性状聚类
研究表明,第Ⅰ类群包含80份材料,占供试材料的29.0%,该类群主要特征为株高较高,单株穗数最多,在四大类群中位居第1位,穗长居中,每穗粒数最少,千粒重最低,多为多穗少粒型种质资源。
第Ⅱ类群包含35份材料,占供试材料的12.7%,该类群主要特征为株高最低,单株穗数较多,每穗粒数较低,千粒重较高,穗长较长,多为矮杆大粒型种质资源。
第Ⅲ类群包含78份材料,占供试材料的28.3%,该类群主要特征为株高和千粒重最高,穗长最长,在四大类群中位居第1位,单株穗数较少,每穗粒数居中,多为高杆高千粒重型种质资源。
表3 供试大麦4种类群材料5个数量性状的平均值差异比较
Tab.3
| 种质类群 Germplasms groups | 种质数量 Germplasms quantitative(份) | 株高 Plant height (cm) | 千粒重 Thousand grain weight (g) | 单株穗数 Pericles per plant | 穗长 Panicle length (cm) | 每穗粒数 Grain per spike |
|---|---|---|---|---|---|---|
| Ⅰ | 80 | 89.21 | 48.34 | 4.22 | 7.89 | 20.75 |
| Ⅱ | 35 | 79.23 | 53.91 | 4.09 | 7.94 | 20.92 |
| Ⅲ | 78 | 89.65 | 54.27 | 3.30 | 8.12 | 21.54 |
| Ⅳ | 83 | 84.01 | 50.71 | 3.21 | 5.56 | 42.43 |
图1
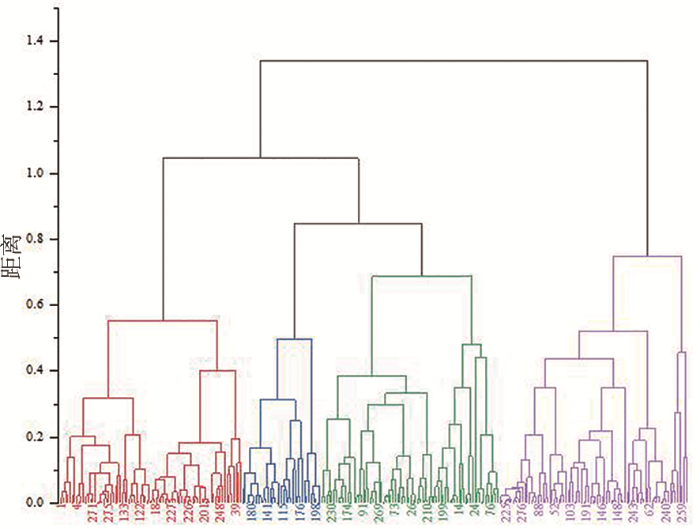
图1
供试材料基于5个数量性状的聚类
Fig.1
The clustering results of the test materials based on 5 quantitative traits
2.4 大麦种质资源品质性状聚类
研究表明,第Ⅰ类群包含144份材料,占供试材料的52.17%,该类群主要特征为水分和淀粉含量最高,在三大类群中位居第1位,粗蛋白和粗纤维含量最低。
第Ⅱ类群包含98份材料,占供试材料的35.51%,该类群主要特征为粗纤维含量最高,淀粉含量最低,水分和粗蛋白含量居中。
表4 供试大麦3种类群材料4个品质性状的平均值差异比较
Tab.4
| 种质类群 Germplasms groups | 种质数量 Germplasms quantitative(份) | 水分 Water content (%) | 粗蛋白湿基 Crude protein (%) | 粗纤维湿基 Coarse fiber (%) | 淀粉湿基 Starch (%) |
|---|---|---|---|---|---|
| Ⅰ | 144 | 9.80 | 11.29 | 3.03 | 56.55 |
| Ⅱ | 98 | 9.57 | 11.84 | 3.98 | 54.22 |
| Ⅲ | 34 | 9.50 | 12.92 | 3.43 | 55.30 |
图2
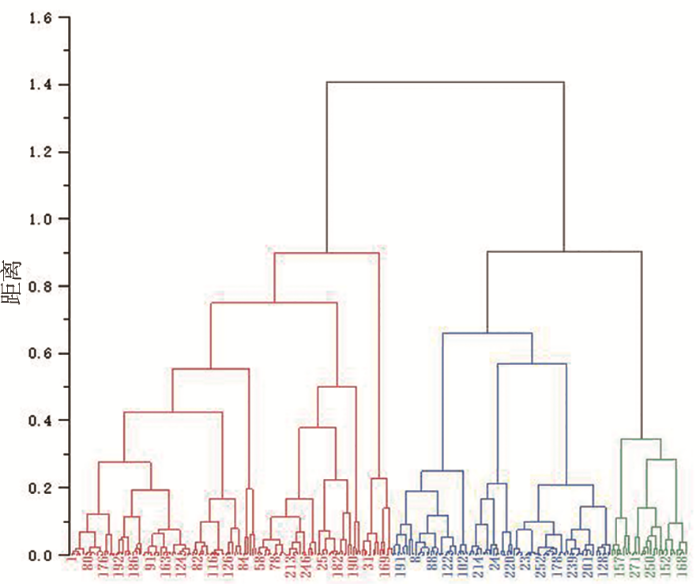
图2
供试材料基于4个品质性状的聚类
Fig.2
The clustering results of the test materials based on 4 quantitative traits
2.5 基于数量性状和品质性状的大麦种质资源聚类
研究表明,数量性状第Ⅳ类群为高产群体,品质性状第Ⅲ类群为优质群体。
数量性状第Ⅰ类群包含80份材料,其中对应的品质性状第Ⅰ类群、第Ⅱ类群、第Ⅲ类群分别为41、26、13份种质材料。
数量性状第Ⅱ类群包含35份材料,其中对应的品质性状第Ⅰ类群、第Ⅱ类群、第Ⅲ类群分别为24、3、8份种质材料。
数量性状第Ⅲ类群包含78份材料,其中对应的品质性状第Ⅰ类群、第Ⅱ类群、第Ⅲ类群分别为49、18、11份种质材料。
数量性状第Ⅳ类群包含83份材料,其中对应的品质性状第Ⅰ类群、第Ⅱ类群、分别为30、51份种质材料。
第Ⅲ类群高产群体中品质性状为优质的种质材料仅有2份,分别为2013-7-DM073和吉引2013-7-DM-105。表5
表5 供试大麦基于5个数量性状和4个品质性状的聚类
Tab.5
| 数量性状种质 Quantitative trait germplasm grop | 品质性状种质类群Ⅰ Quality character germplasm group I | 品质性状种质类群Ⅱ Quality character germplasm groupⅡ | 品质性状种质类群Ⅲ Quality character germplasm groupⅢ |
|---|---|---|---|
| 数量性状 种质类群Ⅰ Quantitative trait germplasm group Ⅰ | 2013-7-DM034、2013-7-DM048、2013-7-DM050、2013-7-DM060、2013-7-DM067、2013-7-DM071、2013-7-DM072、2013-7-DM092、2013-7-DM107、2013塔引004、2013塔引005、2013塔引006、2013塔引013、2013塔引022、2013塔引023、吉引2013-7-DM-121、吉引DM2012-15、吉引DM2012-18、吉引DM2012-22、吉引DM2012-25、吉引DM2012-28、哈引ASTN DM2012-3、吉引2013-7-DM-002、吉引2013-7-DM-003、吉引2013-7-DM-005、吉引2013-7-DM-017、吉引2013-7-DM-024、吉引2013-7-DM-034、吉引2013-7-DM-037、吉引2013-7-DM-044、吉引2013-7-DM-049、吉引2013-7-DM-063、吉引2013-7-DM-066、吉引2013-7-DM-076、吉引2013-7-DM-085、吉引2013-7-DM-092、吉引2013-7-DM-098、吉引2013-7-DM-100、2013-7-Y003、2013-7-Y002、2013-7-DM007 | 2013-7-DM043、2013-7-DM045、2013-7-DM046、2013-7-DM049、2013-7-DM051、2013-7-DM053、2013-7-DM062、2013-7-DM069、2013-7-DM077、2013-7-DM084、吉引2013-7-DM-114、吉引DM2012-4、吉引DM2012-5、吉引DM2012-10、吉引DM2012-20、哈引ASTN DM2012-7、吉引2013-7-DM-009、吉引2013-7-DM-035、吉引2013-7-DM-059、吉引2013-7-DM-065、吉引2013-7-DM-067、吉引2013-7-DM-073、吉引2013-7-DM-082、吉引2013-7-DM-091、吉引2013-7-DM-095、吉引2013-7-DM-096 | 2013-7-DM037、吉引2013-7-DM-106、吉引2013-7-DM-11、吉引DM2012-16、吉引DM2012-35、吉引DM2012-40、哈引ASTN DM2012-1、吉引2013-5-DM-001、吉引2013-7-DM-025、吉引2013-7-DM-055、吉引2013-7-DM-080、吉引2013-7-DM-094、2013-7-DM003 |
| 数量性状 种质类群Ⅱ Quantitative trait germplasm group Ⅱ | 2013-7-DM036、2013-7-DM087、2013-7-DM093、2013塔引002、2013塔引017、吉引2013-7-DM-113、吉引2013-7-DM-120、吉引2013-7-DM-122、吉引DM2012-8、吉引DM2012-9、吉引DM2012-13、吉引DM2012-19、吉引DM2012-26、哈引ASTN DM2012-5、塔引DM2012-1、吉引2013-7-DM-006、吉引2013-7-DM-012、吉引2013-7-DM-031、吉引2013-7-DM-050、吉引2013-7-DM-052、吉引2013-7-DM-088、吉引2013-7-DM-101、吉引2013-7-DM-102、2013-7-Y013 | 吉引2013-7-DM-118、吉引DM2012-21、2013-7-Y005 | 2013-7-DM089、吉引DM2012-1、吉引DM2012-14、吉引DM2012-23、吉引DM2012-24、哈引ASTN DM2012-2、哈引ASTN DM2012-8、吉引2013-7-DM-010 |
| 数量性状 种质类群Ⅲ Quantitative trait germplasm group Ⅲ | 2013-7-DM052、2013-7-DM058、2013-7-DM059、2013-7-DM068、2013-7-DM070、2013-7-DM074、2013-7-DM075、2013-7-DM076、2013-7-DM078、2013-7-DM079、2013-7-DM081、2013-7-DM083、2013-7-DM091、2013-7-DM097、2013-7-DM099、2013-7-DM108、2013-7-DM109、2013塔引007、2013塔引008、2013塔引011、2013塔引014、2013塔引015、2013塔引020、吉引2013-7-DM-108、吉引2013-7-DM-112、吉引DM2012-7、吉引DM2012-34、吉引DM2012-42、哈引ASTN DM2012-4、吉引2013-7-DM-016、吉引2013-7-DM-018、吉引2013-7-DM-019、吉引2013-7-DM-020、吉引2013-7-DM-021、吉引2013-7-DM-026、吉引2013-7-DM-027、吉引2013-7-DM-028、吉引2013-7-DM-029、吉引2013-7-DM-041、吉引2013-7-DM-042、吉引2013-7-DM-043、吉引2013-7-DM-045、吉引2013-7-DM-046、吉引2013-7-DM-062、吉引2013-7-DM-070、2013-7-Y012、2013-7-Y009、2013-7-DM002、2013-7-DM004 | 2013-7-DM044、2013-7-DM047、2013-7-DM055、2013-7-DM056、2013-7-DM057、2013-7-DM100、2013塔引001、吉引DM2012-6、吉引DM2012-11、吉引2013-7-DM-014、吉引2013-7-DM-032、吉引2013-7-DM-057、吉引2013-7-DM-071、吉引2013-7-DM-072、吉引2013-7-DM-074、吉引2013-7-DM-081、吉引2013-7-DM-084、吉引2013-7-DM-097 | 2013-7-DM039、2013-7-DM042、2013-7-DM080、2013-7-DM106、吉引DM2012-27、吉引DM2012-32、吉引DM2012-36、吉引DM2012-41、吉引DM2012-44、吉引2013-7-DM-004、吉引2013-7-DM-078 |
| 数量性状 种质类群Ⅳ Quantitative trait germplasm group Ⅳ | 2013-7-DM061、2013-7-DM063、2013-7-DM064、2013-7-DM066、2013-7-DM082、2013-7-DM085、2013-7-DM096、2013-7-DM103、2013-7-DM104、2013塔引003、2013塔引009、2013塔引010、2013塔引016、2013塔引018、吉引2013-7-DM-110、吉引2013-7-DM-115、吉引DM2012-3、吉引DM2012-12、吉引DM2012-38、吉引DM2012-39、哈引ASTN DM2012-6、哈引ASTN DM2012-9、吉引2013-7-DM-007、吉引2013-7-DM-011、吉引2013-7-DM-038、吉引2013-7-DM-064、吉引2013-7-DM-090、吉引2013-7-DM-093、吉引2013-7-DM-099、2013-7-Y010 | 2013-7-DM035、2013-7-DM038、2013-7-DM040、2013-7-DM041、2013-7-DM054、2013-7-DM065、2013-7-DM086、2013-7-DM088、2013-7-DM090、2013-7-DM094、2013-7-DM095、2013-7-DM098、2013-7-DM101、2013-7-DM102、2013-7-DM105、2013塔引012、2013塔引019、2013塔引021、2013塔引024、2013塔引025、吉引2013-7-DM-104、吉引2013-7-DM-107、吉引2013-7-DM-109、吉引DM2012-2、吉引DM2012-17、吉引DM2012-29、吉引DM2012-30、吉引DM2012-31、吉引DM2012-33、吉引DM2012-37、吉引2013-7-DM-008、吉引2013-7-DM-015、吉引2013-7-DM-023、吉引2013-7-DM-040、吉引2013-7-DM-047、吉引2013-7-DM-051、吉引2013-7-DM-054、吉引2013-7-DM-056、吉引2013-7-DM-058、吉引2013-7-DM-061、吉引2013-7-DM-075、吉引2013-7-DM-083、吉引2013-7-DM-087、吉引2013-7-DM-089、吉引2013-7-DM-103、2013-7-Y004、2013-7-Y006、2013-7-Y008、2013-7-DM005、2013-7-DM006、2013-7-DM008 | 2013-7-DM073、吉引2013-7-DM-105 |
3 讨论
遗传多样性是开展基因挖掘和品质改良的前提,对广泛收集的种质进行遗传多样性分析,对资源各农艺性状进行全面的鉴定和评价[17]。大麦的品种选育方式以种间杂交选育为主,收集和筛选丰富的大麦种质资源是提高大麦产量、品质和抗性的必要前提[18]。农艺性状是作物评价的直观依据,与作物的产量和品质相关,对其遗传多样性的分析是丰富作物种质资源类型、指导作物亲本选配、进行作物育种的基础[19]。徐肖等[20]对86份青藏裸大麦的主要农艺性状的表型多样性进行分析,发现8个质量性状和4个数量性状均存在着丰富的多样性,并且数量性状遗传多样性高于质量性状。夏腾飞等[21]对267份青稞种质的9个数量性状进行研究表明,各性状的遗传多样性指数相近,平均值为2.03,存在丰富的表型多样性。田朋佳等[22]对西藏不同地区的140 份大麦13个农艺性状进行遗传多样性分析发现,数量性状的遗传多样性普遍较高,变幅是1.96~3,农艺性状变异较大,其中分蘖数的遗传多样性指数最大为3,变异系数( 29.8% )。蔡羽等[3]对来源于全国不同地区的102份库存高秆大麦的15 个表型性状的表型多样性进行分析,发现来源于不同生态区的种质表型多样性不同,结果表明中国库存大麦种质资源具有丰富的表型多样性,为后续开展遗传多样性分析和优异种质挖掘奠定了基础。试验研究对大麦的7个质量性状进行分析,发现存在较为广泛的遗传多样性,5个数量性状遗传多样性指数较接近,平均值为1.92,并且数量性状的遗传多样性高于质量性状。研究结果同上述结果相一致,说明供试大麦资源具有较高的丰富度和均匀度,遗传多样性广泛,并且数量性状的变异较质量性状更为丰富,可能与研究材料来源于不同的生源地、且不同地区生态环境差异大有关。聚类分析在作物品种资源上的应用已逐步深入,虽然其分析结果因分析的样本数及方法等不同有所不同,但其在一定程度上反映品种间的差异,为品种资源的分析研究及育种亲本的选择提供重要的理论依据[23]。
4 结论
将供试大麦276份种质划分为4大类群,第Ⅰ类群(包括80份材料)表现为每穗粒数少、千粒重较低、单株穗数较多资源;第Ⅱ类群(包括35份材料)属于矮杆大粒型资源;第Ⅲ类群(包括78份材料)属于高杆多穗长穗型资源;第Ⅳ类群(包括83份材料)属于少穗多粒型资源。各类群大麦的农艺性状表现各有优势。第Ⅲ类群高产群体中品质性状为优质的种质材料仅有2份,分别为2013-7-DM073和吉引2013-7-DM-105。
参考文献
日本不同棱型大麦种质资源农艺性状的差异
[J].
Study on germplasm resource among different rowed barley
[J].
大麦品质和农艺性状的通径分析
[J].
Path analysis of quality and agronomic characters in barley germplasm
[J].
大麦表型多样性分析及优异饲草种质资源筛选
[J].
为探究我国库存大麦种质资源的表型多样性及其变异规律,同时挑选高生物产量的大麦遗传资源发展饲草大麦,本研究以来源于全国不同地区的102份库存高秆大麦种质为材料,通过Shannon-Weaver多样性指数、相关分析、聚类分析、主成分分析等方法对15个表型性状进行综合评价。结果表明:15个性状中拔节期底部叶片黄化、倒伏性、灌浆期底部叶片黄化、千粒重变异较大,粒宽、株高变异较小,表型多样性指数范围为0.729 - 0.957,皮裸性、千粒重、籽粒周长、灌浆期株高等性状的遗传多样性较高。不同来源种质间表型多样性比较结果表明,冬大麦生态区表型多样性(0.721)高于西南及青藏高原裸大麦生态区(0.706)。相关分析、主成分分析、聚类分析均表明,皮裸性是影响大麦种质表型差异的重要影响因素,而倒伏、拔节期和灌浆期底部叶片黄化等饲草相关性状相对独立。聚类分析将102份种质分为4类,相同来源地的种质聚在一起。本研究结果表明我国库存大麦种质资源具有丰富的表型多样性,从中筛选的高秆、大叶、抗倒伏等优异性状的大麦资源,为后续培育青饲、青贮专用型大麦品种提供了资源保障。
Characterization of phenotypic variation in cultivated barley provided elite genetic germplasm with potential breeding for silage barley
[J].Cultivated barley (Hordeum vulgare L. ssp. vulgare) is one of oldest domesticated crops and four most important cereal species. In addition to end-use for animal feeding, malting and human diets using barley grains, the straw of cultivated barley serves as a silage grass for animals in some parts of barley cultivation areas (e.g. Tibet). In order to identify elite barley resources in future breeding for silage barley varieties, this study analyzed the phenotypic variation 102 barley germplasm accessions from National Crop Genebank of China in Chongqing, south-west part of China. We determined the phenotypic variation on 15 traits and conducted statistical analysis using Shannon-Weaver Index, correlation analysis, cluster analysis and PCA analysis. These results suggested that the Shannon-Weaver Index (H’) ranged from 0.729 to 0.957. The higher phenotypic variation was detected on leaf yellowing at heading and filling stages, plant lodging and 1000-grain weight, but the narrowed diversity was found on grain width and plant height. In addition, we found that the phenotypic diversity of winter barley ecological region (0.721) was slightly higher than that of southwest and Tibet Plateau region (0.706). Four subpopulations were clarified by clustering analysis, being consistent with the geographic origins of tested materials. Moreover, a number of barley germplasm resources, which showed high plant height, lodging resistance, large leaves, staying green of old leaves, were identified. These elite barley germplasms might become useful potentially in breeding for silage barley varieties in areas of Chongqing, China.
不同大麦品种(系)营养功能成分差异比较
[J].
Determination of functional compositions in different barley cultivars(lines) by spectrophotometry
[J].
大麦亲本材料农艺性状鉴定及遗传多样性分析
[J].
Identification of agronomic traits and genetic diversity analysis of barley parent materials
[J].
西藏部分青稞主要农艺性状分析
[J].
Analysis of main agronomic traits of several Tibetan naked barley
[J].
大麦种质资源表型性状遗传差异分析
[J].【目的】研究大麦种质资源遗传多样性,提高大麦种质资源利用效率,为新疆大麦品种选育和品种改良提供参考依据。【方法】以157份国内外大麦品种(系)为材料,分析14个主要农艺性状进行遗传多样性。【结果】6个质量性状的遗传多样性指数变幅为0.18~0.65,平均值为0.41,穗姿和株型多样性较丰富;8个数量性状的遗传多样性指数变幅为1.37~2.04,变异系数差为4.07%~24.69%,平均值为1.89,表型多样性丰富;8个质量性状之间相互影响,彼此关联;前3个特征值的主成分,累计贡献率达73.351%。【结论】157份材料分成4大类群,第Ⅰ类为高千粒重型资源,第Ⅱ类为矮杆大粒型资源,第Ⅲ类高秆多穗长穗型资源,第Ⅳ类表现为少穗多粒型资源。
Genetic diversity analysis of main agronomic characters of barley germplasm resources
[J].【Objective】 Understanding and mastering the genetic diversity of germplasm resources is beneficial to improving the utilization efficiency of barley germplasm resources in this region.【Methods】 In this study, 157 domestic and foreign barley germplasm resources were used as materials to analyze the genetic diversity of its 14 main agronomic traits. 【Results】 6 qualitative traits had a wide range of genetic diversity indexes, ranging from 0.18 to 0.65, with an average of 0.41. The diversity of ear pose and plant type was abundant; the coefficients of variation of 8 quantitative traits had obvious differences, and the range of variation was 0.41. The range of genetic diversity index was 4.07%-24.69%, the range of genetic diversity index was 1.37-2.04, and the average value was 1.89. There was rich phenotypic diversity; the correlation analysis results showed that the 8 quality traits influenced each other and were related to each other; The principal components of the first 3 eigenvalues were selected for analysis, and the cumulative contribution rate was 73.351%. 【Conclusion】 Cluster analysis divides 157 materials into 4 major groups, the first type is high-thousand-grain heavy resources, and the second type is short-stem large-grain resources. Type III resources belongs to high stalks, multiple ears and long ears, and type IV resources has fewer ears and more grains.
基于SSR标记的大麦种质资源遗传多样性分析
[J].
Assessment of the genetic diversity of barley germplasm resources based on SSR markers
[J].
新疆大麦品种(系)不同生态区适应性鉴定试验
[J].
Assessment of the adaptability of barley varieties(lines) to different ecological zones in Xinjiang
[J].
世界大麦种质资源遗传多样性分析
[J].
On genetic diversity among barley germplasm in the world
[J].
143份大麦种质资源主要农艺性状遗传多样性分析
[J].
Genetic diversity of main agronomic characters of 143 barley germplasm resources
[J].
44份西藏大麦材料农艺性状的聚类分析
[J].
Cluster analysis for agronomic traits of 44 Tibetan barley materials
[J].
不同来源大麦对条纹病抗性鉴定及遗传多样性分析
[J].为了解不同来源大麦对条纹病的抗性及遗传多样性,本研究采用三明治法通过人工接种大麦条纹病菌对91份大麦材料进行抗性评价,并通过31个SSR 标记对91份大麦亲本材料进行遗传多样性和群体结构分析。结果表明,人工接种大麦条纹病菌后共鉴定出4份免疫、6份高抗、33份抗病、42份感病和6份高感大麦亲本材料;31对SSR标记共检测出等位基因238个,平均每对标记可检测到7.677个等位基因,等位基因的变幅为2~19;主基因频率变化范围为0.236~0.951,平均值为0.394;基因多样性指数的变幅为0.094~0.871,平均值为0.667;多态性信息量变幅为0.091~0.860,平均值为0.613;遗传相似系数变异范围为0.103~1.000,平均值为0.522;在GS值为0.783水平上可将参试材料聚为3个大类群,各大类分别包含86、2和3份材料;群体遗传结构分析表明,供试大麦亲本材料分为3个亚群,每个亚群分别包含47、33和32份材料,且在91份材料中,Q>0.6的材料占总数的97.80%。本研究经抗病鉴定及分子标记结果综合分析,可为挑选抗病亲本辅助抗大麦条纹病优良品种的选育提供参考。
Resistance identification and genetic diversity analysis of barley genotypes from different sources to barley stripe disease
[J].In order to detect the resistance genes of germplasm resources against barley stripe disease and understand their genetic diversity, in this study 91 barley accessions were inoculated with barley stripe disease pathogen by sandwich method, followed by the genetic diversity analysis using 31 polymorphic SSR markers. Of them, four accessions were identified to be complete resistance (free of diseased symptom), 6 highly resistant, 33 resistant, 42 susceptible and 6 high susceptible. Genotyping by 31 SSR markers disclosed 238 alleles, with a mean of 7.677 alleles in each of markers and the range of 2 to 19 alleles. The frequency of major loci ranged from 0.236 to 0.951, with an average of 0.394. The gene diversity index ranged from 0.094 to 0.871, with an average of 0.667. PIC ranged from 0.091 to 0.860, with an average of 0.613, while GS ranged from 0.103 to 1.000, with an average of 0.522. This collection were divided into three groups at the genetic similarity coefficient level of 0.783, each containing 86, 2 and 3 accessions, respectively. The population structure analysis suggested three groups, each including 47, 33 and 32 materials, respectively. 97.80% of 91 genotypes had Q value greater than 0.6. Gained from disease resistance identification and genotyping, this study provided reference for selecting parental lines in breeding of barley leaf stripe resistant varieties.
111份多棱大麦种质主要农艺性状的遗传多样性
[J].
Analysis of the genetic diversity of the main agronomic traits of 111 multi-rowed barley germplasm accessions
[J].
裸大麦种质资源研究进展
[J].
Research progress in naked barley germplasm resources
[J].
大麦引进种质资源表型的多样性与模糊聚类分析
[J].
Phenotypic diversity analysis and fuzzy clustering in barley germplasm resources introduced from abroad
[J].
青藏高原青稞及其他地区大麦种子表型的多样性分析
[J].
Phenotypic diversity analysis of seed traits in barley germplasm from Qinghai-Tibetan Plateau and other regions
[J].
不同地区大麦品种农艺性状比较与西藏青稞品种改良
[J].
Comparison on agronomic traits of barley from different regions and improvement of Tibet naked barley
[J].
青藏高原裸大麦种质资源形态多样性分析
[J].为了解青藏高原裸大麦区种质资源的遗传多样性,提高该地区裸大麦种质资源利用效率,对86份青藏裸大麦种质资源的形态多样性进行鉴定。结果表明:该地区裸大麦种质资源遗传多样性丰富,主穗长的形态多样性指数最高,为2.05;株型的遗传多样性指数最低,为0.47。总体表现为数量性状形态多样性指数大于质量性状。基于形态性状的聚类分析把86份材料聚为4类,第Ⅰ类是二棱种质,为高秆大穗大粒型,第Ⅱ类种质为矮秆多粒长穗型,第Ⅲ类种质为短穗型,第Ⅳ类种质为高秆多粒型。采用主成分分析进行种质评价,提取前7个主成分即可概括全部性状的大部分生物学信息,筛选出适合江苏地区生长的优良种质资源,为裸大麦种质资源的收集和利用提供理论依据。
Morphological diversity of hulless barley accessions from Qinghai-Tibetan Plateau
[J].To study the genetic diversity of hulless barley accessions from Qinghai-Tibetan Plateau and improve their utilization efficiency, the morphological diversity of 86 hulless barley accessions from Qinghai-Tibetan Plateau was identified in this study. The results showed that genetic diversity of hulless barley accessions was ample, morphological diversity index of main panicle length was the highest with the value of 2.05. Plant type had the lowest genetic diversity index with the value of 0.47. In general, genetic morphological index of quantitative traits were greater than those of the quality traits. The tested materials were divided into four groups by cluster analysis. Barley accessions in group Ⅰ were two-rowed, high plant height, big spike and big grain. Barley accessions in group Ⅱwere short plant height, multi grain and long spike. Barley accessions in group Ⅲ were short panicle. Barley accessions in group Ⅳ were high plant height and multi grain. Based on the principal components, comprehensive evaluation of various materials was carried out, seven principal components were extracted to summarize most of the biological information of all traits. Excellent germplasm resources suitable for growth in Jiangsu Province were screened out. This study provided a theoretical basis for the collection and utilization of hulless barley accessions.
267份青藏高原青稞种质材料的表型多样性分析
[J].
The genotypic diversity analysis of 267 six-rowed hulless barley accessions from the Qinghai-Tibetan Plateau
[J].
140份西藏大麦种质资源遗传多样性分析
[J].
Genetic diversity analysis of 140 barley germplasm resources in Tibet
[J].

{kind=link}
{kind=link}
{kind=link}
{kind=link}